Science. 2019 Jan 4;363(6422):eaav1483. doi: 10.1126/science.aav1483. Epub 2018 Dec 13.
Synaptotagmin-3 drives AMPA receptor endocytosis, depression of synapse strength, and forgetting (シナプトタグミン-3がAMPA受容体のエンドサイトーシスを駆動しシナプスの強度を低下させ、忘却を促す)
Ankit Awasthi # 1, Binu Ramachandran # 1, Saheeb Ahmed 1, Eva Benito 2 3, Yo Shinoda 1, Noam Nitzan 1, Alina Heukamp 1, Sabine Rannio 1, Henrik Martens 4, Jonas Barth 2 3, Katja Burk 1, Yu Tian Wang 5, Andre Fischer 2 3, Camin Dean 6
Affiliations
1Trans-synaptic Signaling Group, European Neuroscience Institute, 37077 Goettingen, Germany.
2German Center for Neurodegenerative Disease, 37075 Goettingen, Germany.
3Department of Psychiatry and Psychotherapy, University Medical Center Goettingen, 37075 Goettingen, Germany.
4Synaptic Systems GmbH, 37079 Goettingen, Germany.
5Brain Research Center and Department of Medicine, University of British Columbia, Vancouver, BC V6T2B5, Canada.
6Trans-synaptic Signaling Group, European Neuroscience Institute, 37077 Goettingen, Germany. c.dean@eni-g.de. #Contributed equally.
忘却と受容体の除去
シナプス後膜表面へのAMPA受容体の出入りは、シナプスの強度を調節し、学習と記憶の根底にある。Awasthiらは、膜タンパク質であるsynaptotagmin-3(Syt3; シナプトタグミン3)がニューロンのシナプス後膜のエンドサイトゾーンに多く存在し、AMPA受容体の内在化を促進することを発見した(Perspective by Mandelberg and Tsienを参照)。シナプトタグミン3を過剰発現またはノックダウンした神経細胞では、シナプス伝達と短期可塑性は変化しなかった。しかし、Syt3ノックアウトマウスの神経細胞では、シナプスの長期抑圧が消失し、長期増強が減衰していることがわかった。Syt3ノックアウトマウスでは、空間学習は変化しなかったが、水迷路の空間記憶課題では、忘却と再学習に障害の徴候が見られた。
サイエンス、今号p.eaav1483; p.31も参照。
Structural Abstract
〇はじめに
記憶は、脳内の分子や細胞の変化として保存される。シナプス(神経細胞間の接続部位)は、神経細胞間の通信の有効性を調整する能力により、記憶を保存することができる。このシナプス可塑性という特性により、脳は記憶を保存し、取り出すことが可能である。忘却とは、記憶を潜在化させたり、有用性を低下させたりすることで、記憶を取り出せなくすることである。しかし、記憶が忘れられるかどうかは、どのようなメカニズムで決まるのだろうか。そのメカニズムとして、シナプス後細胞膜上の神経伝達物質受容体数の調節が挙げられる。この受容体は、シナプス前に放出された神経伝達物質を電気信号に変換することで、シナプス伝達を媒介します。神経細胞の活動は、受容体を挿入することでシナプスを強化し、シナプス後膜から受容体を除去することでシナプスを弱体化させます。受容体の輸送は、神経細胞へのカルシウムの流入によって制御されるが、この制御を媒介するカルシウムセンサーは知られていない。
〇解説
シナプトタグミンは、カルシウムを感知して膜融合を誘発する。神経細胞の培養を刺激すると、ほとんどのシナプトタグミンのアイソフォームがカルシウムを介し細胞膜にドッキングするが、シナプトタグミン3(Syt3)はシナプス後膜に存在する。AMPA型あるいはNMDA(N-メチルd-アスパラギン酸)受容体を刺激すると、AMPA受容体(脳内の高速シナプス伝達の大部分を仲介する)とSyt3の内在化が誘導される。このことから、Syt3が活動によって誘導された受容体の内在化を仲介して、シナプスを弱め、忘却を引き起こすという興味深い可能性が浮上した。我々は、シナプトタグミン3(Syt3)をアイソフォーム特異的抗体で画像化し、脳切片と神経細胞培養の電気生理学的手法を用いて受容体輸送におけるSyt3の役割を検証し、マウスの空間記憶課題を用いて忘却におけるSyt3の役割を検証した。
〇結果
Syt3は、シナプス後膜のエンドサイトゾーン(シナプス後の密度に近いクラスリンに富む領域)に存在する。Syt3はGluA2 AMPA受容体サブユニットと結合し、クラスリンを介したエンドサイトーシスによるAMPA受容体の活性依存的な内在化に関与する2つのタンパク質であるAP2およびBRAG2とも結合する。Syt3は、基本的なAMPA受容体の輸送には影響を与えない。しかし、Syt3をノックアウトするか、カルシウム結合を欠損させたSyt3を発現させると、AMPA、NMDA、あるいは電気生理学的刺激によるシナプス強度の長期抑制によって誘導されるAMPA受容体の内部化が阻害される。また、シナプスの強さの長期増強が通常減衰するAMPA受容体の内在化も阻害する。これらの効果は、Tat-GluA2-3Yペプチドの急性投与によって野生型でも再現される。このペプチドは、GluA2の細胞質尾部にあるチロシンリッチ(3Y)モチーフへのSyt3の結合を競合的に阻害する。空間記憶課題において、Syt3をノックアウトしたマウス(Syt3ノックアウトマウス)は、逃避位置を正常に学習するが、以前に学習した位置に固執する。これは、以前に獲得した記憶を忘れることができないためと説明することができる。野生型マウスにTat-GluA2-3Yペプチドを注射すると、空間記憶の忘却の欠如を模倣し、この効果はSyt3ノックアウトマウスで消滅する。
〇結論
記憶の持続と劣化は、十分に理解されていない分子機構によって支配されている。我々は、カルシウムを介したAMPA受容体の内在化を誘発し、シナプス伝達の弱体化と空間記憶の忘却をもたらすシナプトタグミンアイソフォームを発見し、マウスでそのメカニズムを解明した。
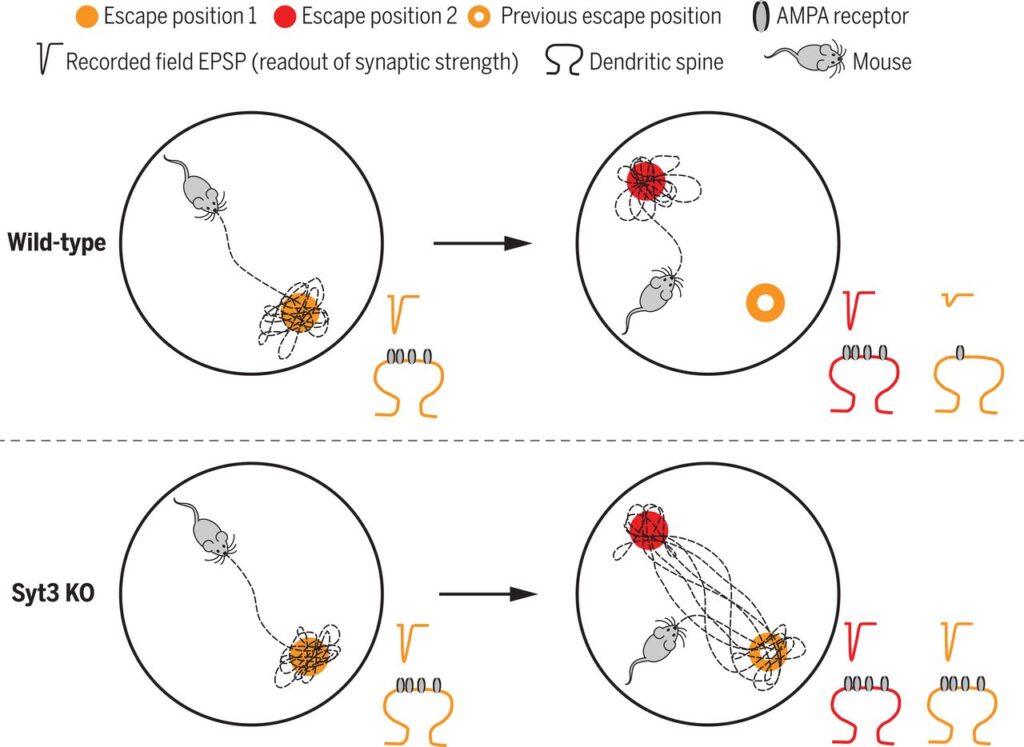
Syt3ノックアウトマウスは忘却しない。
野生型マウスもSyt3ノックアウトマウスも水迷路で脱出位置を覚えることができるが、このとき対応するシナプスはAMPA受容体の増加により強化される。これらのシナプスは、例えば新しい脱出位置を学習したときなど、記憶が不要になると受容体が除去され、弱くなる。Syt3ノックアウトマウスは受容体を除去できないので、以前の脱出位置を忘れることができない。
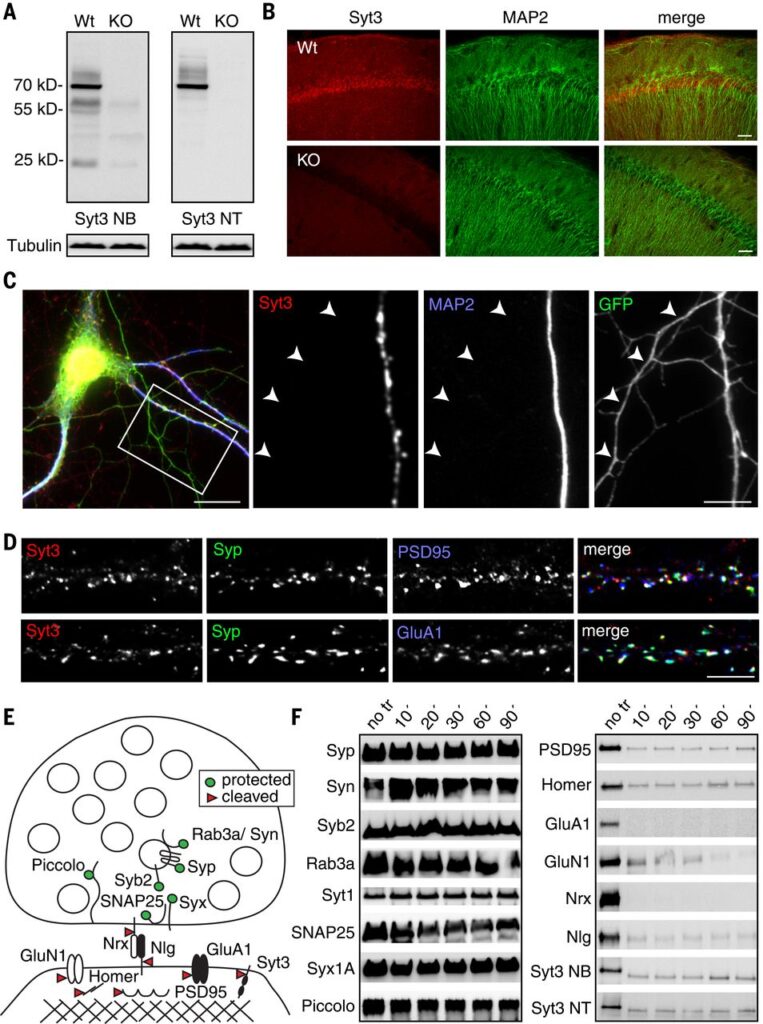
Fig. 1 Syt3 is postsynaptic.
(A) Syt3 antibodies recognize a specific band of the expected molecular weight in wild-type (WT) but not in Syt3 knockout (KO) mouse brain homogenates in Western blots. Syt3 NB is a monoclonal antibody from Neuromab, and Syt3 NT is a polyclonal antibody developed by Synaptic Systems. Tubulin is a loading control. (B) Syt3 immunoreactivity is detected on pyramidal cell bodies and dendrites in the CA1 region of hippocampal slices but not in Syt3 knockouts. MAP2 marks dendrites. Scale bar, 50 μm. (C) Syt3 is predominantly in dendrites in hippocampal neurons transfected with GFP and immunostained with MAP2 to mark dendrites (MAP2-positive) or axons (GFP-positive/MAP2-negative) (n = 20 neurons/3 cultures). Scale bar, 20 μm (left), 5 μm (right). (D) Syt3 localizes to synapses marked by synaptophysin and PSD95 (top) or GluA1 (bottom) in hippocampal cultures. Scale bar, 5 μm. (E) Schematic of a synaptosome. The presynaptic side reseals, whereas the postsynaptic side does not, leaving postsynaptic proteins accessible to trypsin cleavage. (F) Presynaptic proteins are protected from trypsin cleavage, whereas postsynaptic proteins (including Syt3) are cleaved.
Discussion
筆者らは、活動依存的なAMPA受容体の内在化、LTD、LTPの減衰、および空間記憶の忘却にSyt3が必要であることを発見した。レスキュー実験では、LTDとLTPの減衰には、シナプス後のSyt3によるカルシウム感知が必要であった。GluA2-3Yペプチドは、GluA2受容体尾部の3Y領域へのSyt3の結合を競合的に阻害する。このペプチドを野生型に導入すると、Syt3ノックアウトの表現型であるAMPA受容体の活性依存的な内在化の欠如、LTD、LTPの減衰、忘却を模倣した。Syt3ノックアウトマウスでは、このペプチドの効果は消失し、Syt3がGluA2-3Yに依存したAMPA受容体の内在化機構に関与していることが示唆された。
このデータから、シナプス後のエンドサイトゾーンに存在するSyt3が、カルシウムのない状態でAP-2とBRAG2に結合しているというモデルが生まれた。そして、GluA2は、神経活動時のカルシウムの増加に応じてSyt3と結合し、エンドサイトゾーンに蓄積される可能性がある。これにより、GluA2はBRAG2に近接する可能性があり、一過性の相互作用によりBRAG2とArf6が活性化され、クラスリンやAP-2を介して受容体のエンドサイトーシスを促進することができる。PICK1はAMPA受容体のエンドサイトーシスにも重要であることから、Syt3とPICK1の相互作用が問題となる。2つの可能なメカニズムが考えられる。PICK1はエンドサイトーシスの後、受容体を内部に封じ込め、Syt3の下流で作用する可能性がある。あるいは、PICK1は刺激後に一過性にGluA2、そしてAP-2と結合し、エンドサイトーシスゾーンにAMPA受容体を集め、その後の内在化を促進する可能性もある。したがって、PICK1がSyt3の上流で、あるいはSyt3と協調して、GluA2をエンドサイトゾーンに導くという可能性もある。
最近、Syt1/Syt7のシナプス後発現を阻害すると、LTPは消失するがLTDには影響がないことが報告された。このように、異なるSytがシナプス後膜から受容体を挿入したり除去したりして、それぞれLTPとLTDを媒介する可能性がある。Sytはエキソサイトーシスとエンドサイトーシスの両方を制御することができるが、そのカルシウム親和性によってどちらか一方に偏ることになる。エンドサイトーシスは、エキソサイトーシスよりも遅い時間スケールで起こる。カルシウム濃度が低下すると、Syt3のような高親和性の細胞は活性を維持するが、Syt1のような低親和性の細胞は不活性化する。あるいは、エンドサイトーシスを起こしやすいシトは、Ca2+との結合を拡張するタンパク質複合体と相互作用したり、静止状態でタンパク質と結合し、Ca2+の結合により放出されてエンドサイトーシスを起こす可能性もある
記憶力はしばしば記憶の最も重要な側面とみなされるが、忘却も同様に重要である。忘却の欠陥は深刻な結果をもたらし、例えば、心的外傷後ストレス障害につながることがある。筆者らのデータは、Syt3が、AMPA受容体が内在化され、LTDとLTPの減衰を促進する忘却メカニズムに重要な分子であることを明らかにしている。