本日は、医学類M5のTさんが研究進捗報告を行いました。いつも通り、美しくまとめられたスライドでした。セミナーを聴きながら、Tさんを実習で試問した時の知識の多さに驚いたことを思い出しました。もう一人のM5学生の輝きももう少し近くで見てみたいと思っています。フロンティア医科学学位プログラムの大学院生が下記の論文紹介を行いました。
Nature Communications, 2025年
論文番号16:918。
投稿日は2023年11月20日、受理日は2025年1月15日、オンライン公開日2025年1月22日
Nonapoptotic caspase-3 guides C1q-dependent synaptic phagocytosis by microglia アポトーシス非依存性カスパーゼ-3は、ミクログリアによるC1q依存性シナプス貪食を誘導する
Megsumi Andoh, Ryuta Koyama et al. Department of Translational Neurobiology, National Institute of Neuroscience, National Center of Neurology and Psychiatry, Kodaira, Tokyo 187-8502, Japan.
Abstract
カスパーゼは、脳発達過程における神経細胞のアポトーシスを媒介することが知られている。しかし本研究では、プレシナプスにおけるカスパーゼ-3の非アポトーシス性活性化が、ミクログリアによるシナプス貪食を駆動することを示す。
新たに確立したマウス由来の培養系において、シナプスのカスパーゼ-3をリアルタイムで観察し、時空間的に操作したところ、神経活動の増加がプレシナプスに局所的なカスパーゼ-3活性化を引き起こし、それによって補体によるシナプスのタグ付けが促進されることが明らかになった。
高解像度ライブイメージングにより、カスパーゼ-3活性化は、軸索の切断を伴うことなく、シナプス選択的かつ補体依存的なミクログリア貪食を促進することが示された。
さらに、抑制性プレシナプスにおける活動依存的なカスパーゼ-3活性化は、マウスにおいてミクログリアによる貪食を誘導し、発作感受性を高めた。この感受性の増加は、ミクログリアの補体受容体を遺伝的に欠損させることで元に戻った。
したがって、局所的で非アポトーシス性のカスパーゼ活性は、補体依存的なミクログリアによるシナプス貪食を誘導し、神経回路を再構築する。
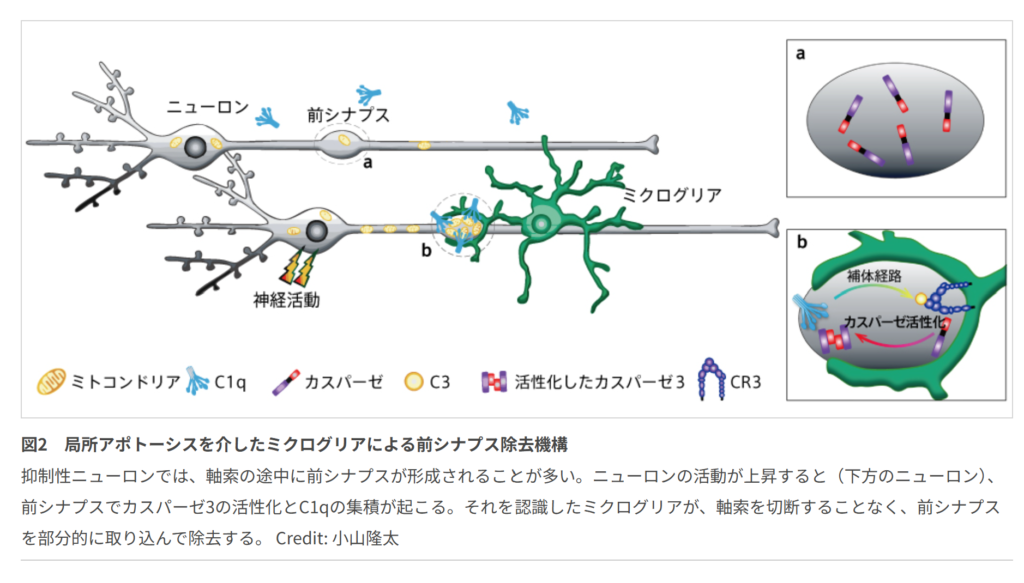
ミクログリアがシナプスを食べる瞬間を可視化する
方法
本研究では、主に神経細胞・ミクログリア・アストロサイトを含むマウス由来の神経グリア共培養系と、マウス熱性けいれんモデルを用いている。in vitroでは、神経活動を高めるために hM3Dq DREADD を導入し、CNO(clozapine-N-oxide)20 μM を投与しました。プレシナプスは synaptophysin-mCherry で標識され、切断型カスパーゼ-3、C1q、C3、ホスファチジルセリンなどの局在が免疫染色やライブイメージングで解析された。
カスパーゼ-3活性のリアルタイム検出には、FRET型プローブ synaptophysin-mSCAT3 が用いられました。このプローブでは、カスパーゼ-3活性化により DEVD 配列が切断され、mECFP/mVenus比 が上昇する。著者らは、mECFP/mVenus比が 1以上 のプレシナプスをカスパーゼ-3活性化部位として扱っている。カスパーゼ阻害には、カスパーゼ-3阻害薬 Z-DEVD-FMK 10 μM、カスパーゼ-9阻害薬 Z-LEHD-FMK 20 μM が用いられた。また、ミトコンドリア経路の関与を調べるために、Baxチャネル阻害薬 2 μM、Apaf-1阻害薬 NS3694 2 μM も使用されている。in vivoでは、P11マウス に熱性けいれんを誘導し、歯状回中分子層の興奮性シナプス、抑制性シナプス、補体タグ付け、ミクログリア貪食を解析しました。熱性けいれん後の発作感受性は、3 mg/kg のカイニン酸投与により評価された。
Results
最初に、神経活動の亢進がプレシナプスで局所的なカスパーゼ-3活性化を引き起こすことが示された。CNO投与 6時間後、synaptophysin-mCherry陽性プレシナプスに切断型カスパーゼ-3シグナルが増加し、この増加は Z-DEVD-FMK 10 μM により抑制された。Fig.1では、CNOにより切断型カスパーゼ-3体積が有意に増加し、DMSOとの比較で p < 0.0001、CNO + DEVDとの比較で p = 0.0007 が示されている。
次に、CNO投与により、プレシナプスで C1qA-647 の局在が増加した。Fig.2では、C1qA + CNO条件でプレシナプスへのC1q局在が上昇し、C1qA + DMSOとの比較で p = 0.0076 が示されている。さらに、CNOとC1qAが同時に存在する条件で、ミクログリアによるプレシナプス貪食が最も増加した。ライブイメージングでは、C1qA + CNO条件で貪食されたプレシナプス数が増え、p = 0.0288 と示されている。
Fig.3では、光遺伝学的にカスパーゼ-9/カスパーゼ-3を局所的に活性化する CRY2-casp9 系が用いられた。青色光刺激により、刺激されたプレシナプスだけでなく、同じ軸索上の他のプレシナプスにも切断型カスパーゼ-3が観察された。C1q存在下で青色光刺激を行うと、プレシナプスへのC1q局在が増加し、カスパーゼ-9阻害薬 Z-LEHD-FMK により抑制された。これにより、カスパーゼ-3活性化自体がC1qタグ付けの十分条件であることが示された。
in vivoの熱性けいれんモデルでは、Fig.4において、熱性けいれん後 1時間 では興奮性神経細胞マーカー Reelin 陽性細胞のc-Fos発現が増加したが、4時間後 には対照レベルに戻った。一方、抑制性神経細胞マーカー GAD67 陽性細胞では、c-Fos発現が 4時間後 も上昇し続けた。さらに、熱性けいれん 6時間後 には、切断型カスパーゼ-3が興奮性シナプスマーカー VGLUT1 ではなく、抑制性シナプスマーカー VGAT と強く共局在した。
Fig.5では、熱性けいれん 6時間後 に C1q と C3 の点状シグナル密度が上昇した。C1qでは p = 0.0007、C3では p = 0.0056 が示されている。また、VGAT陽性抑制性シナプスで、切断型カスパーゼ-3、ホスファチジルセリン、C1q、C3の共局在が増加した。この変化は、Z-DEVD-FMKを熱性けいれん誘導 15分前 に投与することで抑制された。
Fig.6では、熱性けいれん後にミクログリア側の補体受容体 CR3 が増加した。CR3 mRNAは熱性けいれん後 1時間 および 4時間 で上昇し、CR3タンパク質シグナルも 6時間後 に増加した。ミクログリアによる貪食は、興奮性シナプス VGLUT1 では有意に変化せず、p = 0.7907 でしたが、抑制性シナプス VGAT では大きく増加し、p < 0.0001 でした。
さらに、熱性けいれん後 24時間 および 72時間 では、興奮性シナプス密度は安定していた一方、抑制性シナプス密度のみが低下した。CR3ノックアウトマウスでは、熱性けいれん後の抑制性シナプス貪食増加と抑制性シナプス密度低下が消失した。最後に、熱性けいれん 3日後 にカイニン酸を投与すると、野生型マウスでは発作スコアが上昇したが、CR3ノックアウトマウスではこの増加が抑えられた。
Discussion
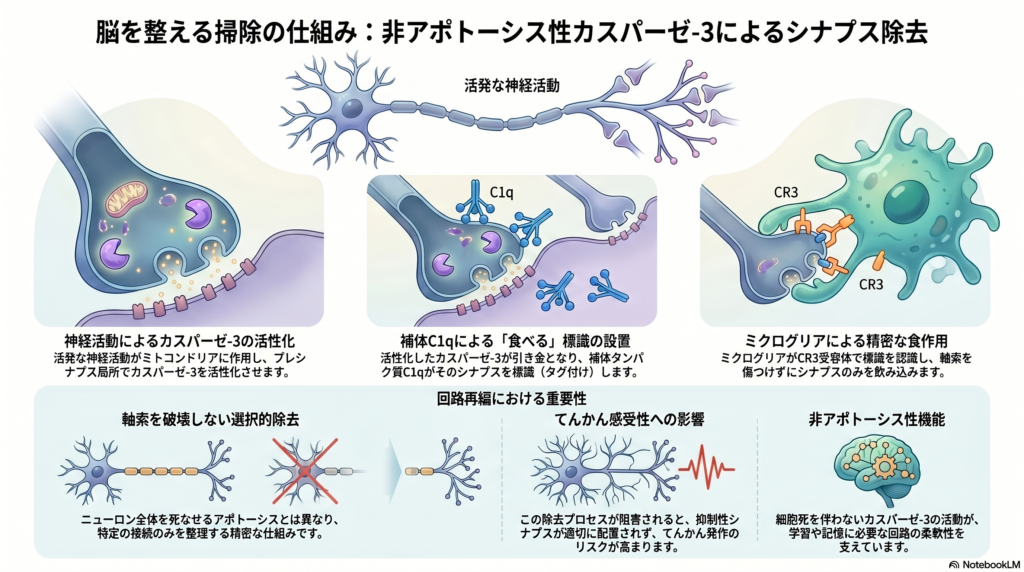
本研究の議論では、神経活動の亢進がプレシナプスにおいて局所的な非アポトーシス性カスパーゼ-3活性化を引き起こし、それがC1qによる補体タグ付けを誘導し、最終的にミクログリアによるシナプス選択的貪食につながるという新しい機構が示された。従来、カスパーゼ-3は主に神経細胞死を実行する因子として理解されてきたが、本研究では細胞死を伴わず、シナプス単位で局所的に働く分子としての役割が明確にされたのである。特に、ミクログリアが軸索を切断せずにプレシナプスのみを貪食する様子が観察された点は、従来の貪食とは異なる精密なシナプス再編成機構の存在を示す重要な結果である。
著者らは、カスパーゼ-3活性化が補体タグ付けを促す仕組みとして、ホスファチジルセリンの外在化がC1q結合を助ける可能性を挙げている。これによりC3を含む補体カスケードが進み、ミクログリアの補体受容体CR3が標識されたシナプスを認識して貪食を開始するという流れである。また、カスパーゼ-3活性化がプレシナプスに限局する理由として、プレシナプスに存在するミトコンドリア、カルシウム流入、シトクロムc放出、IAPsによるアポトーシス抑制などが関与する可能性があると考察されている。さらに、同一軸索上の別のプレシナプスにもカスパーゼ-3活性化が広がることから、モータータンパク質や細胞骨格を介した活性化シグナルの伝播も想定される。
熱性けいれんモデルでは、抑制性シナプスにおいてカスパーゼ-3活性化、C1q/C3タグ付け、ミクログリア貪食が起こり、その後に抑制性シナプス密度が低下した。これは神経回路の抑制性入力を弱め、結果として発作感受性を高める要因になると考えられる。実際、CR3欠損により貪食や抑制性シナプス減少、発作感受性の増加が抑制されたことから、この過程が補体依存的であることが強く支持される。
一方で、本研究には限界もある。in vitroではシナプス選択的貪食を高解像度で観察できたが、in vivoで同様の現象を直接可視化するには技術的制約がある。また、DREADDや熱性けいれんによる神経活動亢進は生理的条件より強い可能性があり、発達期の正常なシナプス剪定や学習依存的可塑性にも同じ機構が働くかは今後の課題である。総じて、本研究はカスパーゼ-3を細胞死因子ではなく、神経活動に応答して補体依存的なシナプス再編成を導く局所制御因子として位置づけた点に大きな意義がある。