Molecular Neurobiology, 2022年 掲載巻号 59:5750–5765
オンライン公開日 2022年7月4日、受理日は2022年6月10日です。DOI 10.1007/s12035-022-02936-x
Tbr1 Misexpression Alters Neuronal Development in the Cerebral Cortex (Tbr1の異所性発現は大脳皮質における神経発達を変化させる)
Inmaculada Crespo et al. Instituto Cajal-Consejo Superior de Investigaciones Científicas (CSIC), Avenida Doctor Arce 37, 28002 Madrid, Spain
Abstract
転写因子(TF)の発現変化は脳発生に極めて重要であり、神経発達障害の背景にもなり得る。実際、T-box brain1(Tbr1)は新皮質第VI層の形成に不可欠な転写因子であり、この遺伝子の変異や微小欠失は、自閉スペクトラム症(ASD)を伴うヒト大脳皮質の形成異常と関連している。興味深いことに、Tbr1の発現上昇もASD様症状の出現と関連づけられているが、新皮質発生中にTbr1量が増加することの影響を扱った研究は限られている。
本研究では、主に第II–IV層ニューロンを産生する時期である胎生14.5日(E14.5)のマウス神経前駆細胞(NPC)において、Tbr1の異所性発現が及ぼす影響を解析した。E18.5までに、細胞は中間帯および大脳皮質深層に蓄積し、一方で上層では少なくなった。これと一致して、第V–VI層におけるSox5陽性細胞の割合は増加し、第II–IV層におけるCux1陽性細胞の割合は減少した。
生後7日(P7)では、移動異常はより軽度になっていたが、上層および深層においてSox5陽性細胞の割合が高い状態は認められた。異常なニューロン移動は、Tbr1異所性発現によって誘導される多極性ニューロンおよび双極性ニューロン形態の変化による部分があると考えられた。また、Tbr1異所性発現は樹状突起の成長と分岐を減少させ、脳梁の形成も障害した。
以上の結果は、皮質神経前駆細胞におけるTbr1の異所性発現が、ニューロン移動、ニューロン分化指定、樹状突起発達、脳梁路形成を遅延または障害することを示している。したがって、発生過程において異所性のTbr1発現上昇を引き起こす遺伝的変化は、大脳皮質の脳形成異常を誘発し得る。
Results
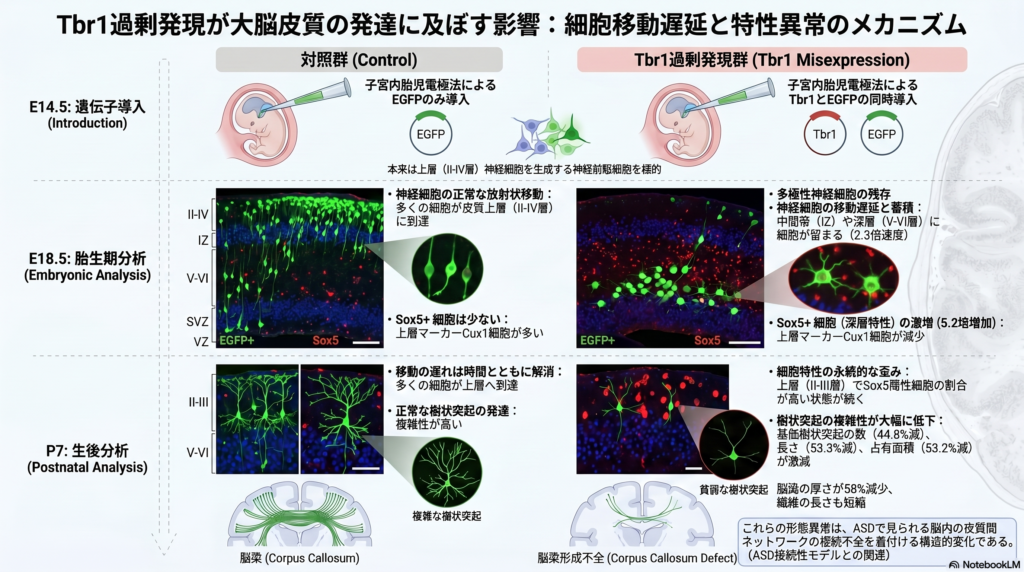
本研究では、E14.5の大脳皮質神経前駆細胞にTbr1を異所性発現させ、その後のニューロン発生への影響を解析した。pCAG-Tbr1-EGFPを導入した群では、GFP陽性細胞のほぼ100%がTbr1を発現していた。一方、対照群ではTbr1とGFPの二重陽性細胞はほとんど見られなかった。EGFP陽性細胞数は、対照群で144.41±11.53、Tbr1群で179.27±24.19であり、導入効率に大きな差はない結果である。
E18.5で細胞の分布を調べると、Tbr1異所性発現によりニューロンの配置が大きく変化していた。中間帯に存在する細胞割合は2.3倍に増加し、上層である第II–IV層の細胞は34.5%減少した。層別解析でも、第VI層のGFP陽性細胞は14.0%±3.26から28.0%±9.38へ、第V層では17.5%±2.64から35.0%±5.71へ増加した。一方、第II–IV層では68.8%±5.79から37.5%±10.72へ減少した。これらの結果は、Tbr1の過剰な発現により、ニューロンが本来到達すべき上層へ移動しにくくなったことを示す。
この異常は、細胞死や前駆細胞増殖の変化では説明しにくい。cleaved caspase-3陽性細胞は両群とも約1%と低く、Ki67陽性細胞やTbr2陽性中間前駆細胞の割合にも有意な差はなかった。つまり、Tbr1異所性発現の主な影響は、細胞の増殖や生存ではなく、ニューロン移動そのものにあると考えられる。
さらに、Tbr1異所性発現はニューロンの形態にも影響した。中間帯の多極性ニューロンは15.93%±1.17から26.60%±0.21へ増加し、第V–VI層の非放射状双極性ニューロンも1.34%±0.43から4.50%±0.65へ増加した。一方、移動に適した放射状双極性ニューロンは98.03%±0.56から93.17%±0.97へ減少した。これは、多極性から放射状双極性への形態転換が乱れたことを示す結果である。
神経細胞の分子的性質にも変化が見られた。Cux1陽性細胞とCTIP2陽性細胞は減少し、Sox5陽性細胞は520%増加した。加えて、第II–III層ニューロンでは樹状突起長が28.4%、樹状突起数が59.2%減少し、脳梁軸索の長さは44.07%、厚さは58.06%減少した。P7でも樹状突起異常とSox5陽性細胞の増加は残っていた。以上より、Tbr1異所性発現は、ニューロン移動、層形成、細胞種指定、樹状突起形成、脳梁形成を広く乱すことが示された。
Discussion
本論文の議論では、Tbr1は大脳皮質発生に必須である一方、その発現量が過剰になっても神経発達を乱す可能性が強調されている。これまでの研究では、TBR1の機能喪失変異や微小欠失がASDや知的障害、大脳皮質形成異常と関連することが主に示されてきた。しかし本研究は、Tbr1の異所性発現、すなわち本来とは異なる時期や細胞での発現上昇も、皮質発生に悪影響を及ぼすことを示した点に意義がある。
著者らは、E14.5において主に第II–IV層ニューロンを産生する神経前駆細胞にTbr1を発現させた。その結果、E18.5では細胞が中間帯や第V–VI層に蓄積し、本来移動すべき第II–IV層へ到達する細胞が減少した。この異常は、細胞増殖、Tbr2陽性前駆細胞数、細胞死の変化では十分に説明できず、ニューロン移動そのものの障害によると考えられている。特に、多極性ニューロンから放射状双極性ニューロンへの形態転換が乱れたことが、放射状グリアに沿った移動の遅延につながった可能性がある。
また、Tbr1異所性発現は細胞の位置だけでなく、神経細胞の分子的アイデンティティにも影響した。上層ニューロンのマーカーであるCux1や、第V層ニューロンに関わるCTIP2が減少し、深層ニューロンに関連するSox5陽性細胞が増加したことから、層形成と細胞種指定の両方が乱れたと解釈される。P7では移動異常は目立ちにくくなるものの、Sox5陽性細胞の増加や樹状突起形態の異常は残存していた。これは、発生初期のTbr1過剰発現が一過性の移動遅延にとどまらず、成熟後の神経回路形成にも影響し得ることを示している。
さらに、樹状突起の成長や分岐、脳梁軸索の伸長も障害されたことから、Tbr1発現上昇は皮質内外の神経接続形成にも影響する可能性がある。これらの所見は、ASDで報告される皮質間結合異常や皮質下異所性灰白質と関連し得る。著者らは、本研究が患者の遺伝子変異を直接再現したモデルではないという限界を認めつつも、Tbr1の発現量は時期的・空間的に厳密に制御される必要があり、その破綻がASDや知的障害に伴う大脳皮質形成異常の一因となる可能性を示したと結論づけている。








