医療科学類4年生が研究進捗報告を、医学類新医学主専攻の学生が論文紹介を行いました。セミナー後に、医学類5年生がフロンティア医科学学位プログラム大学院生に解析の方法を教えました。本日は、某実習は1-5限に実施でしたので、スタッフは疲弊しました。明日も1限から実習+大事なイベントなのでがんばります。
Science Advances 2025年7月2日公開
LHX2 regulates dendritic morphogenesis in layer II/III neurons of the neocortex (LHX2は新皮質II/III層ニューロンの樹状突起形態形成を制御する)
Mahima Bose et al.
Department of Biological Sciences, Tata Institute of Fundamental Research, Mumbai, India
Abstract
哺乳類の新皮質では、左右半球は脳梁を介して相互に情報を伝達している。本研究では、脳梁投射ニューロンの樹状突起アーバーおよびスパイン形成を制御する分子機構を解析した。転写因子LIM Homeodomain 2(Lhx2)は、皮質発生の重要な制御因子であり、postmitoticなII/III層ニューロンおよびその前駆細胞に発現している。前駆細胞あるいはpostmitotic neuronのいずれかでLhx2を欠失させると、共通点を持ちながらも異なる表現型が生じ、樹状突起アーバーの縮小、スパイン形態の異常、および電気生理学的特性の変化が認められた。形態学的異常は前駆細胞でLhx2を欠失させた場合により重度であり、この表現型はbasal progenitor特異的なLhx2欠失によっても再現された。さらに、前駆細胞でのLhx2欠失は、その後生じるpostmitotic neuronにおいてNeurog2を異常に上昇させ、このNeurog2のノックダウンにより表現型は部分的に回復した。また、Lhx2をいずれの段階で欠失させた場合にも、Wntシグナル伝達経路関連遺伝子の発現上昇が認められた。加えて、postmitotic neuronにおけるβ-CATENINの恒常的活性化は、Lhx2 postmitotic mutantで観察された樹状突起およびスパイン形態異常を再現した。これらの結果は、LHX2依存的な新規樹状突起形成機構を明らかにするとともに、新皮質発生においてLHX2が時間依存的かつ多面的な役割を果たすことを示している。
Results
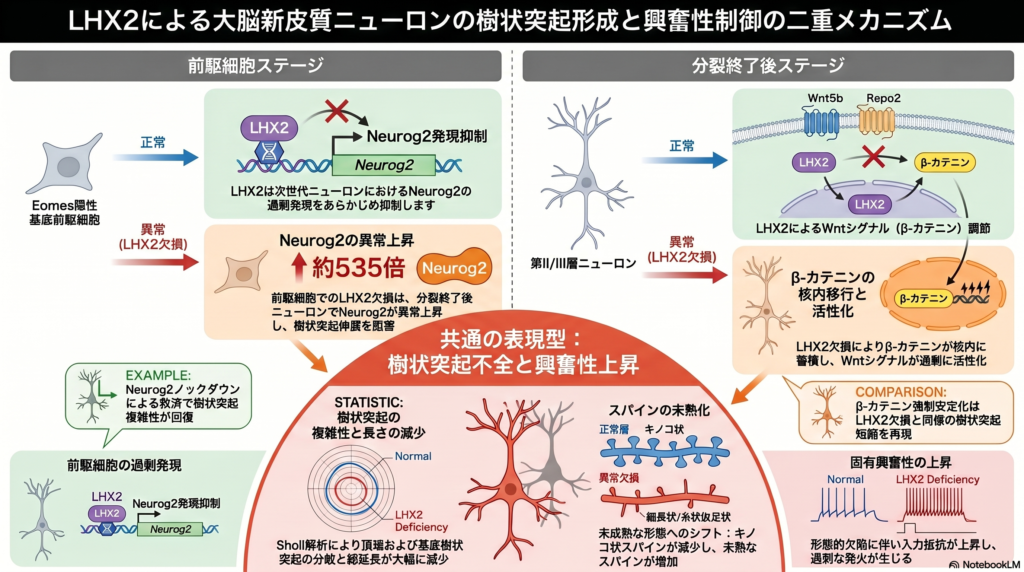
本研究では、転写因子LHX2が大脳新皮質II/III層ニューロンの樹状突起形成およびスパイン成熟に果たす役割を解析した。著者らはE15.5マウス胎仔にin utero electroporationを行い、Lhx2を神経前駆細胞段階(Pg mutant)あるいは出生直後のpostmitotic neuron段階(Pm mutant)で条件的に欠失させ、P30時点で形態学的・電気生理学的解析を実施した。
まず、Neurolucida解析により、Pg mutantおよびPm mutantの両群で樹状突起アーバーが著しく縮小していることが示された。特にapical dendriteおよびbasal dendriteの総長、basal dendrite平均長が有意に低下し、Sholl解析では樹状突起分枝複雑性の低下が確認された。これらの異常はPm mutantよりPg mutantでより重度であった。さらに、樹状突起スパイン解析では、成熟型であるmushroom spineが減少し、未成熟型であるthin spineが増加していたことから、Lhx2欠失によってスパイン成熟障害が起こることが示唆された。
次にpatch clamp解析を行ったところ、Pg mutantではinput resistanceが増加し、Pg/Pm両群で発火頻度の増加が認められた。さらに低電流刺激でdepolarization blockが生じるなど、ニューロン興奮性が亢進していた。一方で、これらのニューロンはPOU3F2など上層ニューロンマーカーを保持し、正常にII/III層へ移動し、脳梁投射も維持していたことから、LHX2はニューロンサブタイプ決定ではなく、主に樹状突起形成と機能成熟を制御していることが示された。
RNA-seq解析では、Pg mutantでNeurog2が535倍に異常上昇していた。single-cell RNA-seq解析により、Neurog2はEomes陽性basal progenitorに特異的に発現することが示され、Eomes-Creを用いたbasal progenitor特異的Lhx2欠失でもPg mutantと同様の樹状突起異常が再現された。さらにNeurog2 shRNAを導入すると、apical dendrite異常はほぼ完全に、basal dendrite異常は部分的に rescueされ、スパイン異常も改善した。これより、LHX2はbasal progenitorでNeurog2を抑制することで、後の樹状突起形成を制御していることが示された。
加えて、Pg/Pm mutantではWntシグナル関連遺伝子(Wnt5b、Rspo2、Sfrp1など)が共通して変動していた。Pm mutantでは核内β-catenin蓄積が増加しており、β-catenin gain-of-functionマウスではLhx2 mutantと類似した樹状突起およびスパイン異常が再現された。以上より、LHX2はNeurog2抑制とWnt/β-catenin経路制御を介して、II/III層ニューロンの樹状突起形成と機能成熟を統合的に制御していることが明らかとなった。
Discussion
本研究の議論では、LHX2が神経発生の異なる時期において異なる分子機構を介しながら、新皮質II/III層ニューロンの成熟を制御していることが強調されている。特に著者らは、Lhx2を前駆細胞段階で欠失させた場合と、postmitotic neuron段階で欠失させた場合で類似した樹状突起異常が生じる一方、その分子背景は部分的に異なることを示した。
最も重要な知見の1つは、前駆細胞段階でのLhx2欠失によりNeurog2が異常持続発現する点である。通常、Neurog2はbasal progenitorで一過性に発現し、ニューロンがpostmitoticになる頃には抑制される。しかしLhx2欠失ではこの抑制が破綻し、成熟ニューロンにおいてもNeurog2発現が持続していた。Neurog2はニューロン分化や移動、樹状突起形成に関与する転写因子として知られているが、本研究ではその異常持続発現が樹状突起形成障害の原因となることが示された。実際、Neurog2 knockdownによってapical dendrite異常が完全に rescueされ、basal dendrite異常も部分的に改善した。この結果から、LHX2はbasal progenitor段階でNeurog2発現を適切に抑制することにより、将来のpostmitotic neuronの形態成熟を制御していると考察している。
一方、postmitotic neuron特異的Lhx2欠失ではNeurog2異常は認められなかった。このことから、LHX2のクロマチン制御機能は発生段階によって異なる可能性が示唆された。著者らは、LHX2がNuRD chromatin remodeling complexと相互作用する既報を踏まえ、Lhx2欠失によってNeurog2遺伝子座のepigenetic repressionが維持できなくなる可能性を議論している。
さらに著者らは、Pg/Pm両変異体で共通してWnt signaling関連遺伝子が変動していたことに注目している。特にpostmitotic mutantでは核内β-cateninが増加しており、β-catenin gain-of-functionによりLhx2 mutantと類似した樹状突起・スパイン異常が再現された。これより、LHX2はpostmitotic neuronにおいてcanonical Wnt signalingを抑制することで樹状突起複雑性を維持している可能性が示された。ただし、Wnt signalingを直接抑制してLhx2 mutantを rescueする実験は未実施であり、今後の課題として挙げられている。
また本研究では、Lhx2欠失によりニューロン興奮性が亢進することも示された。これは樹状突起縮小によるinput resistance増加や、イオンチャネル発現変化の可能性を示唆しているが、その詳細な分子機構は未解明である。
最後に著者らは、LHX2 haploinsufficiencyがヒト自閉スペクトラム症、知的障害、小頭症と関連することを踏まえ、本研究が神経発達障害における樹状突起病態理解に重要な知見を与えると結論づけている。








