Cell Reports Volume 30, Issue 10, 10 March 2020, Pages 3492-3505.e5
Layer 6b Is Driven by Intracortical Long-Range Projection Neurons (第6b層は皮質内の長距離投射ニューロンによって駆動される)
TimothyA. Zolnik 1, Julia Ledderose 12, Maria Toumazou 1, Thorsten Trimbuch 2, Tess Oram 3, Christian Rosenmund 2, Britta J. Eickholt 2, Robert N.S. Sachdev 1, Matthew E. Larkum 14
1Institute for Biology, Humboldt-Universität zu Berlin, Berlin, Germany2Neuroscience Research Center, Charité-Universitätsmedizin Berlin, Berlin, Germany3Janelia Research Campus, Ashburn, VA, USA (フンボルト大学ベルリン)
ベルリン・フンボルト大学は、1810年言語学者のヴィルヘルム・フォン・フンボルトによってフリードリヒ・ヴィルヘルム大学 (Friedrich-Wilhelms-Universität) として創立されたベルリンで最も古い大学である。東ドイツ支配下でフンボルト大学と改称され、ドイツ再統一後に現称フンボルト大学ベルリン(=通称ベルリン・フンボルト大学)となる。1945年以前は、「ベルリン大学」として名が通り、欧米諸国や日本の近代大学教育に多大なる影響を与えた。ベルリン・フンボルト大学出身/所属のノーベル賞受賞者は55人にのぼる。ノーベル賞受賞者の大学別ランキングでは、世界で13位、ヨーロッパでは、ケンブリッジ大学、オックスフォード大学に続き3位、ドイツ国内においては1位である。過去には、特に相対性理論で有名な理論物理学者、アルベルト・アインシュタインも10年以上に渡り同大学教授として働いていた。(Wikipedia)
Received 29 March 2019, Revised 20 September 2019, Accepted 10 February 2020, Available online 10 March 2020, Version of Record 10 March 2020.
Highlights
•Layer 6b is heavily innervated by cortical neurons and projects to thalamus and cortex
•Unlike other cortical layers, layer 6b receives little or no thalamic input
•Long-range intracortical neurons produce the strongest input to layer 6b
•Layer 6b is most potently inhibited by somatostatin and parvalbumin interneurons
①第6b層は皮質ニューロンに強く支配され、視床と大脳皮質に投射している。
②他の皮質層とは異なり、第6b層には視床からの入力がほとんどない。
③長距離皮質内ニューロンは第6b層への最も強い入力を生み出す。
④第6b層はソマトスタチンとパルバルブミン介在ニューロンによって最も強力に抑制される。
Summary
大脳新皮質の最深層である第6b層(L6b)は、皮質標的や高次視床に投射しており、覚醒促進神経ペプチドであるオレキシン/ヒポクレチンに反応する唯一の層である。これらの特徴から、L6bは脳の状態を強く調節できることが示唆されるが、L6bへの投射とその影響は未知のままである。我々は、狂犬病に基づく逆行性追跡とチャネルロドプシンによる脳切片の回路マッピングを用いて、マウスの一次体性感覚皮質におけるL6bへの入力をex vivoで調べた。その結果、L6bは対側半球を含む皮質内長距離投射ニューロンから最も強い興奮性入力を受けていることがわかった。対照的に、局所皮質内入力と視床皮質内入力は有意に弱かった。さらに、L6bは他の皮質層よりもはるかに少ない視床皮質入力を受けていることが示唆された。L6bはPVとSST介在ニューロンによって最も強く抑制された。この研究から、L6bは長距離の皮質内情報を統合しており、従来の視床皮質ループの一部ではないことが示された。
Keywords
layer 6, layer 6b, cortex, thalamus, long-range
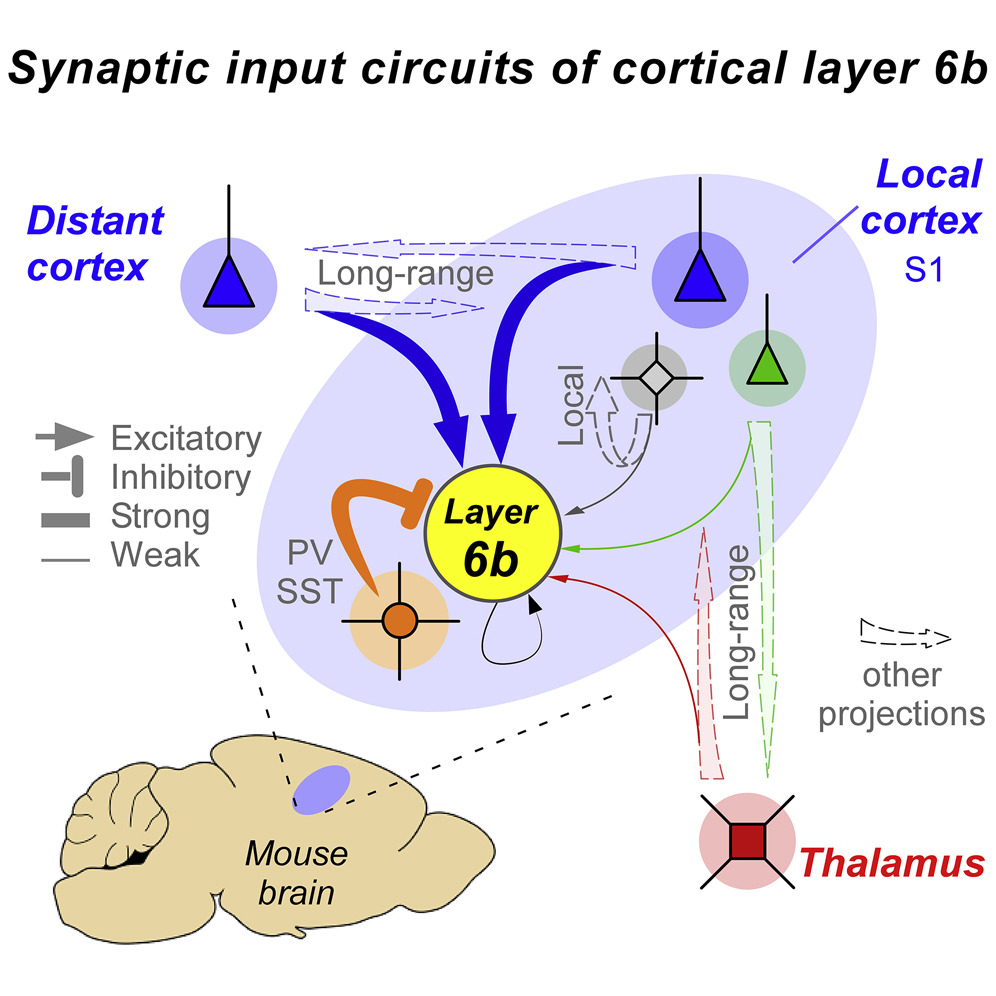
Introduction
100年以上にわたる研究にもかかわらず、大脳新皮質の主要な特徴が未解明であることは注目に値する。大脳新皮質の一番下、灰白質と白質の境界に沿って、6b 層または 7 層と呼ばれる、見落とされがちなニューロンのシートがある(Reep and Goodwin, 1988)。第 6b 層は大脳新皮質の主要な構造の中で最も研究が遅れているものの一つである。
第 6 層(L6)はしばしば単一の均一な層と考えられているが、L6b のニュー ロンは形態学的、遺伝学的、そして発生学的に L6a のニューロンとは異なる。皮質L2-L6aは、亜板から生じるL6bとは異なる前駆体集団に由来し、これらのニューロンは最初に分化し、成熟した生理学的特徴を発現する(Andjelicら、2009、Hoerder-Suabedissen and Molnár、2013、Luskin and Shatz、1985、Marx and Feldmeyer、2013、Marxら、2017、Valverdeら、1989)。このサブプレートには、視床からの入力を受け、感覚刺激に反応する最初の皮質ニューロンが含まれており(Wess et al.
L6bは大脳皮質の中でもユニークな位置にあり、上には新皮質、真下には白質がある。多くのL6bニューロンの樹状突起は白質自体に伸びており、そこにはcallosal軸索(左右両半球をつなぐ)の密なネットワークが横切っている。多くのL6bニューロンの樹状突起はL6を通ってL5にも達しており、そこでは文脈情報(Larkum, 2013)を伝えると考えられているいくつかの長距離経路が接続している(Vandeveldeら, 1996, Watakabe and Hirokawa, 2018, Zhang and Deschênes, 1998)。加えて、視床皮質と皮質視床の軸索もL6bを通過するため、この位置は大脳皮質と視床の間の双方向の情報伝達をモニターするのに適した場所である。
最近の研究では、主にL6aの視床皮質ニューロンが知覚を調節し、皮質回路の利得を制御していることが示されているが(Bortoneら、2014年、Guoら、2017年)、L6bニューロンの行動的・機能的役割はまったく不明である。しかし、L6bのいくつかの特徴は、脳の状態制御と認知に関与していることを示唆している。例えば、感覚皮質において、L6bは覚醒を促進する神経ペプチドであるオレキシン/ヒポクレチンに感受性のある唯一の層であり(Bayerら、2004)、この神経ペプチドが脳内で欠乏すると、睡眠/覚醒障害であるナルコレプシーを引き起こす(Peyronら、2000)。解剖学的には、L6bニューロンは皮質L1(Clancy and Cauller, 1999)、L5a(Viswanathan et al., 2017)、高次視床核(Bourassa et al., 1995, Hoerder-Suabedissen et al., 2018, Killackey and Sherman, 2003, Viswanathan et al. 最近の研究では、サブプレート(L6b)と自閉症や統合失調症を含むいくつかの神経病理との関連も見つかっている(Eastwood and Harrison, 2005, Hoerder-Suabedissen and Molnár, 2013, Nagode et al.)
L6bとの結合を明らかにすることは、L6bニューロンの構造が薄いこと、特異的なマーカーやCreマウス系統がないことなどから、依然として困難であった。本研究では、Creマウス系統を用い、狂犬病に基づく逆行性トレースとチャネルロドプシン支援による脳スライスでの回路マッピングを組み合わせることで、局所的および長距離ネットワークにおけるL6bの解剖学的および機能的統合を調べた。その結果、皮質内投射ニューロンがL6bを強く駆動しているのに対し、視床皮質ニューロンや皮質視床ニューロンのコラテラルはそれほど直接的な影響を受けていないことが明らかになった。これらの結果は、L6bが長距離の皮質内経路からの情報を統合し、視床皮質系からの情報はほとんど統合していないことを示唆している。
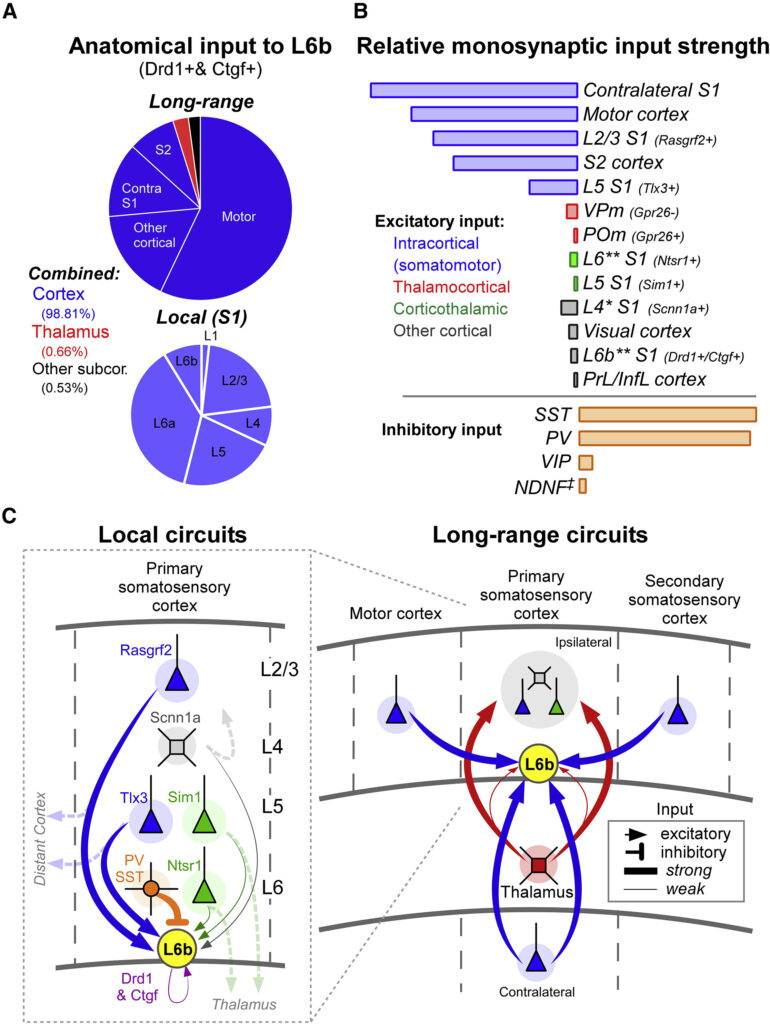
(A) 狂犬病ウイルスを用いたトレーシングに基づく逆行性追跡からの L6b への主な解剖学的入力 (n = 総シナプス前ニューロン 45,694)。 円グラフの各セクションに関連付けられた脳領域にはラベルが付けられ、色分けされている。
(B) テストした主要経路の相対的な直接入力強度 (PSP 振幅 × 応答ニューロンの割合)。 興奮性および抑制性の接続の強さは、そのタイプの他の接続と比較して決まります。 ∗、L6b への強力な間接入力。 ∗∗、L6b 介在ニューロンへの入力は含まれていない。 ‡、もっぱら GABAb 依存性の IPSP。
(C) ローカル (S1) 回線 (左) および長距離回線 (右) 内での L6b の機能統合。 左: 破線の矢印は、各タイプのニューロンの正準投影を示します。
Discussion
本研究は、大脳新皮質の最深層が、局所的な回路と脳全体の回路の中でどのように統合されているかを明らかにすることで、大脳新皮質の理解における重要なギャップを埋めるものである。我々は、一次体性感覚皮質のL6bニューロンへの解剖学的および機能的入力をマッピングし、L6bは視床皮質投射と比較して、長距離皮質内回路から有意に強い入力を受けていることを発見した。
狂犬病ウイルスを用いた神経トレースにより、L6bの長距離シナプス前ニューロンの95%以上が大脳皮質にあることが示された(図2A、7A、7B)。一方、視床には長距離シナプス前ニューロンの3%以下しか存在しなかった。L6bニューロンのサブセットであるDrd1ニューロンとCtgfニューロンのラビートレースを行った(図S1A)。一部の接続が狂犬病によって好まれるかどうかは不明である(Callaway and Luo, 2015)が、本研究では生理学的データが狂犬病データを裏付けた。最も強力で良好に接続されたネットワークには、運動野やS2皮質を含む体性運動領域からの長距離投射が含まれていた。対照的に、視覚野、PrL野、Infl野はL6bを強く駆動しなかった。これらの結果は、S1において、L6bニューロンは主にモダリティに関連した入力を統合していることを示唆している。
対側の体性感覚皮質からの投射は、L6bへの最も実質的な興奮性投射の一つであり、狂犬病トレースは近くのニューロンよりも遠くのニューロンを標識しにくいが、対側の皮質には同側のS2よりも多くのシナプス前ニューロンが含まれていた。この強い対側結合は、L6bが体性感覚系における半球間活動の調整に関与している可能性を示唆している。
S1の長距離投射ニューロンはL6bに強い入力をもたらした。例えば、L2/3とL5のニューロンは、短い局所軸索を持つL4ニューロンからの直接入力よりも強くL6bを駆動した。S1のL5とL6のCTニューロンのローカルコラテラルからL6bへの入力も比較的弱かった。上記の結果と合わせると、このことは、S1内外のニューロンで長距離の皮質内軸索を投射しているニューロンが、L6bへの最も強い入力を生み出していることを示している。
また、L4、L5a、L6aを含むS1ニューロンは、L6bよりも有意に強い視床入力を受けていることもわかった。加えて、これまでの報告では、L1-L6aは相当な視床皮質入力を受けていることが示されている(Audette et al.、2018、Bureau et al.、2006、Constantinople and Bruno、2013、Cruikshank et al.、2010、Cruikshank et al.、2012)。これらの結果を総合すると、L6bは他の層に比べ、視床皮質からの直接的な刺激をほとんど受けていないことが示唆される。ラビートレースはこれらの所見を裏付けている。以前の研究で、様々なタイプのL5錐体ニューロンへのシナプス前ニューロンの少なくとも12%~30%が視床に存在することが示されている(Kim et al. L6bが視床からの直接入力をほとんど受けていないという知見と一致して、in vivo記録では、L6深部ニューロンは他の層と比較して感覚刺激に対して比較的鈍感であることが示されている(de Kockら、2007、Filipkowskiら、2000、Hiraiら、2018、Medini、2011)。実際、入力に関して、L6bとL4は互いに鏡像のようであり、L4は視床皮質ニューロンからの強力な入力を受け、長距離入力はほとんどないのに対し、L6bは視床からの入力を実質的に受けないが、長距離投射の大きなプールからサンプルを得ている。
また、興奮性L6bニューロン間の固有結合のレベルも検証した。ここで我々は、興奮性相互結合は比較的弱いが、興奮性L6bニューロンの光活性化は、しばしば他のL6b興奮性ニューロンにおいて、強固なスパイクを停止させるのに十分な二シナプス抑制を引き起こすことを見いだした(図S5C)。我々はこの抑制がL6のPVおよび/またはSST介在ニューロンから生じている証拠を発見した(図1E、6C-6E)。L6bにおける比較的強い2シナプス抑制は、これがL6bの活動レベルを制御するフィードバック機構であることを示唆している。
シナプス抑制はL6bへの入力に共通する特徴であった。この種の短期可塑性は通常、迅速で信頼性の高いシナプスと関連しており(Sherman and Guillery, 1998)、L6bが長距離情報を迅速に統合できることを示唆している。このことは、L6bが、長距離の皮質内投射ニューロンを介してS1と行き来する文脈情報(Larkum, 2013)を迅速に統合するためのノードであることを示唆している。
〇方法論と比較に関する考察
ここに示した機能的結合の結果は、各投射の入力強度の上限を表している。これにより、それぞれの興奮性投射がどの程度局所回路に関与しているかを調べることができた。例えば、L4はL6bへの直接入力は比較的少ないが、スライス中の他のニューロンを介してL6bへの間接入力は比較的強い。in vitroで作成した縮小標本にはいくつかの利点があるが、in vivoでの各投射からの相対的な影響力は脳の状態によって変化する。とはいえ、我々が報告した皮質内長距離入力の最大(上限)ドライブは、視床皮質入力のそれよりも有意に大きく、したがって、in vivoでは、よほど特殊な状況でない限り、視床皮質投射を上回るだろう。
L6は伝統的な視床皮質ループの重要な構成要素であると考えられているため、L6bへの強い視床皮質駆動がないのは特殊である。VPmとL6のCT(例えばNtsr1)ニューロンはこの相互ループを形成している。しかし、L6bにはVPmからの強い入力がない(図3Dと3K)。さらに、L6bはVPmを強く神経支配していない(Hoerder-Suabedissenら、2018、Viswanathanら、2017)。したがって、L6bは従来の視床皮質ループの外側に位置している。L6aとL6bのもう一つの違いは、後者のみがL1を支配していることである(Clancy and Cauller, 1999)。これらの回路特性は、L6bがL6aとは異なるネットワークに参加していることを示している。とはいえ、これまでの研究で、L6 Ntsr1ニューロンは長距離投射ニューロンからその解剖学的入力の大部分を受け取っていることが示されている(Vélez-Fortら、2014)。また、L6 Ntsr1ニューロンは、皮質内L6aニューロンやL4ニューロン(Crandallら、2017、Wangら、2013)と比べて、視床入力、特に深部Ntsr1ニューロン(Frandoligら、2019)をあまり受けていない。これらの比較に照らすと、Ntsr1ニューロンとDrd1 L6bニューロンの重複は不明であるが、重要な検討事項かもしれない。
〇意義
今回の報告では、L6bが強力な長距離皮質内回路に統合されていることが示された。我々の研究は一次感覚皮質領域で行われたが、L6bは感覚視床経路の影響を劇的に受けなかった。このことは、L6bはせいぜい間接的に感覚処理に関与しているにすぎず、遠く離れた皮質領域からのフィードバックの影響を強く受けていることを示唆している。
いくつかのL6bニューロンは高次視床を強く支配しており、例えばPOm(図S1E;Hoerder-Suabedissenら、2018、Shimaら、2016、Viswanathanら、2017)は運動野やS2野を含む皮質を広く支配している(Hooksら、2013、Ohnoら、2012、Viaeneら、2011a)。POmは運動野とS2皮質の皮質内投射ニューロンを興奮させ、それによって間接的にL6bを興奮させ、さらにL6bがPOmを調節し、全体として大規模な視床-皮質-皮質回路を形成している可能性がある。さらに、L6bニューロンの皮質出力は、L5錐体ニューロンが主要な標的であり、皮質や皮質下の標的に強く広く投射し、脳の状態、行動、知覚に影響を与えるニューロンであることを示唆している。全体として、L6bは長い間無視されてきたし、比較的少数のニューロンを含んでいるという事実にもかかわらず、我々の結果は、大脳皮質の作動を理解する上で極めて重要かもしれないユニークな機能を持っていることを示唆している。
“Journal Club (January 9, 2023)” への3件のフィードバック