Microtubule instability stems from the low energy of tubulin dimer interactions, which sets the growing polymer close to its disassembly conditions. Molecular motors use ATP hydrolysis to produce mechanical work and move on microtubules. This raises the possibility that the mechanical work produced by walking motors can break dimer interactions and trigger microtubule disassembly. We tested this hypothesis by studying the interplay between microtubules and moving molecular motors in vitro. Our results show that molecular motors can remove tubulin dimers from the lattice and rapidly destroy microtubules. We also found that dimer removal by motors was compensated for by the insertion of free tubulin dimers into the microtubule lattice. This self-repair mechanism allows microtubules to survive the damage induced by molecular motors as they move along their tracks. Our study reveals the existence of coupling between the motion of molecular motors and the renewal of the microtubule lattice.
In vitroの研究では、微小管がモーターの層を滑走する際に、プロトフィラメントや二量体を失う可能性があることが示された。しかしこれらの実験では、分子モーターによって生じる機械的力に関するほとんどのin vitro研究と同様に、微小管の解重合を阻止するために微小管が安定化されていた。これらの条件は、成長する微小管末端でチューブリン二量体が取り込まれた後、GTP-チューブリンがGDP-チューブリンに急速に加水分解される生理的条件を反映していない。GDP-チューブリンの格子はGTP-チューブリンの格子よりも安定性が低いため、格子の解体は初期の実験で示唆されたよりも頻繁に起こる可能性がある。そこで我々は、キネシンとダイニンのモーターが微小管に与える機械的な力の影響を、格子がGDP-チューブリンで構成されるような条件下で調べることにした。まず微小管の「グライディング・アッセイ」を用い、小型フローセルのガラス面にモーターを取り付けた。微小管とATPを加えると、活性モーターが生み出す力によって、微小管がモーターでコーティングされた表面を滑るように移動する。
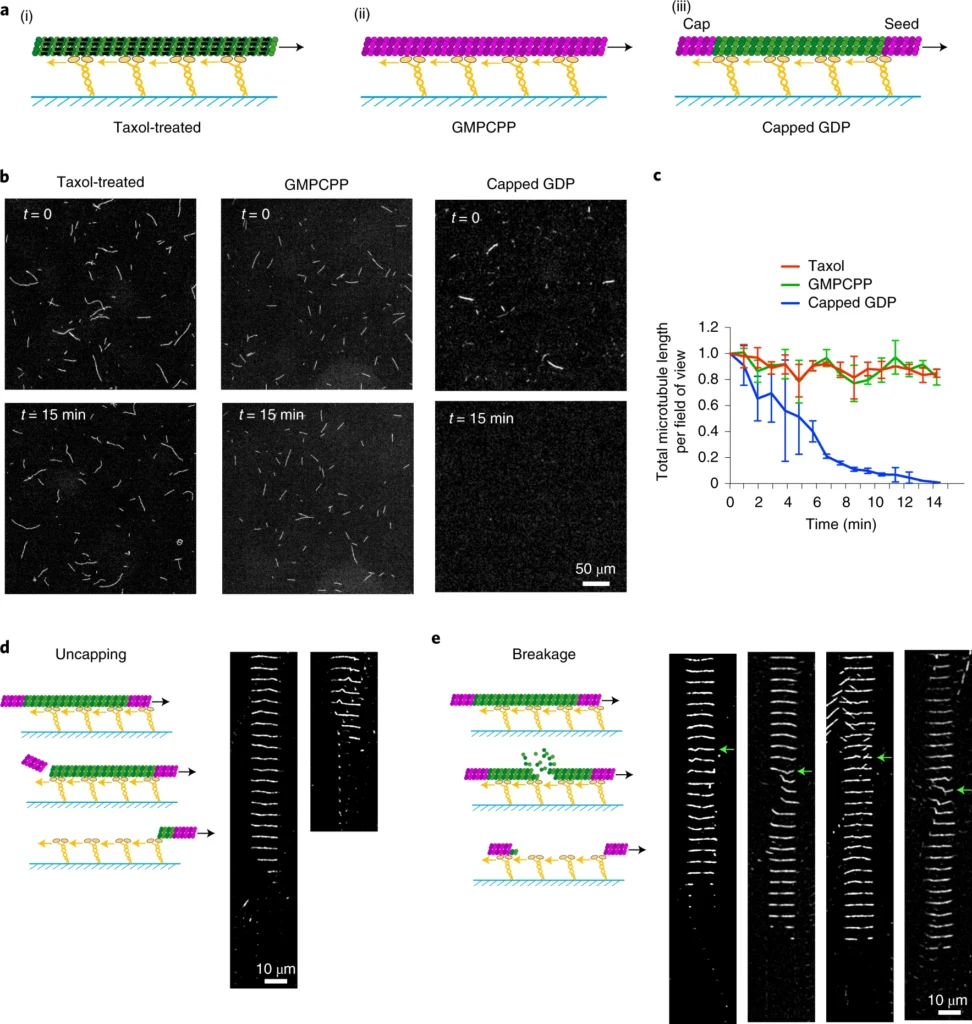
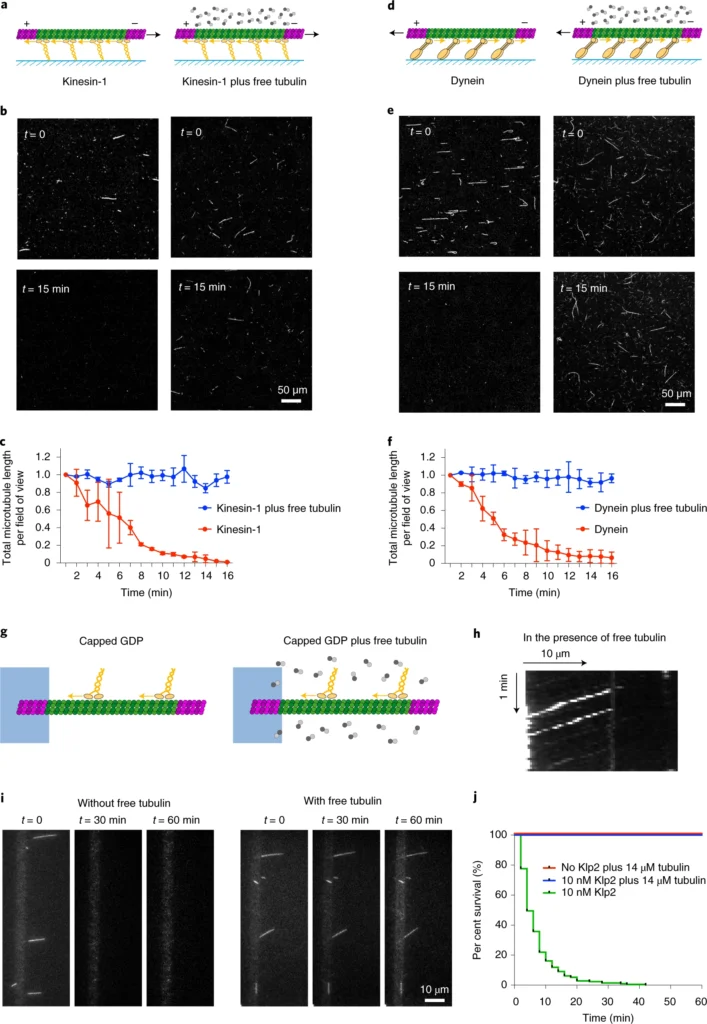
a, Schematic representation of the different types of microtubules used in gliding assays. Taxol-treated (purple) microtubules (i) and GMPCPP-microtubules (magenta dimers; ii) are stabilized along their length. Capped GDP-microtubules (iii) are stabilized at their ends (magenta dimers) but the central part (green dimers) is not stabilized. Molecular motors (yellow) are attached to the cover glass (blue) and propel the microtubules along the glass surface in the presence of ATP. b, Images of gliding microtubules when motors are activated by the addition of ATP (time t = 0, top row) and 15 minutes later (t = 15 min, bottom row) in the three conditions described in a. Scale bar, 50 µm. c, Quantification of microtubule length variations in the experiments shown in b. The microtubule lengths were measured for all microtubules in the 600-µm-wide fields every minute over 15 minutes. Values were normalized with respect to the initial length. Number of independent experiments N = 2; taxol-treated microtubules n = 55; GMPCPP-microtubules n = 42; capped GDP-microtubules n = 42. Error bars represent standard deviation. d, Schematic representation of microtubule disassembly induced by the loss of the cap. Two examples of microtubules that disassembled from an end during the gliding assay are shown. Images are vertical montages from a video where images were acquired every 5 s. Scale bar, 10 µm. e, Schematic representation of microtubule disassembly induced by breakage of the central region of the microtubule. Four examples of microtubules that break and disassemble during the gliding assay are shown. Images are vertical montages from a video where images were acquired every 5 s. Green arrows point to bending events that precede the weakening and eventual breakage of the microtubule. Scale bar, 10 µm.a, Schematic representation of the gliding assay of capped GDP-microtubules on kinesin-1 in the absence (left) or presence (right) of free non-labelled tubulin dimers. b, Images of gliding microtubules when motors are activated by the addition of ATP (t = 0, top row) and 15 minutes later (t = 15 min, bottom row) in the two conditions described in a. Scale bar, 50 μm. c, Quantification of microtubule length variations in the experiments shown in b. The microtubule lengths were measured for all microtubules in the 600-µm-wide fields every minute for 15 minutes. Values were normalized with respect to the initial length. Data in the absence of free tubulin dimers are shown in red and correspond to the data shown in Fig. 1b; data in the presence of 14 µM free dimers are shown in blue. Values were normalized with respect to the initial intensity. Number of independent experiments N = 2, without free tubulin, n = 42; with free tubulin, n = 74; where n is the number of microtubules. Scale bar, 50 μm. Error bars represent standard deviation. d–f, The same images as in a, b and c, but in the presence of yeast dynein instead of kinesin-1. Number of independent experiments N = 2, without free tubulin, n = 24; with free tubulin, n = 12. Error bars represent standard deviation. g, Schematic representation of the motility assay in the absence (left) and presence (right) of free non-labelled tubulin dimers. h, Kymograph showing the position of fluorescent Klp2 along a microtubule over time. The presence of free dimer did not interfere with the ability of the motor to walk towards the microtubule minus end. i, Image sequences show microtubules after the addition of Klp2 motors in the absence (left) or presence (right) of 14 µM free tubulin dimers. Images were taken every 2 min. Scale bar, 10 µm. j, Survival curve of microtubules in the presence of molecular motors and absence of free tubulin (green), in the presence of molecular motors and 14 µM free tubulin dimers (blue) or in the absence of motors and presence of 14 µM free tubulin dimers (red). Data were acquired in the conditions described in i. N = 1; no motor, n = 108; 10 nM Klp2 with free tubulin, n = 70; 10 nM Klp2 without free tubulin, n = 212.
モーターによる微小管の破壊と修復が細胞内で起こるかどうかは、まだ確立されていない。GTP-チューブリンや、CLIP-170(cytoplasmic linker protein of 170 kDa)やCLASP(cytoplasmic linker associated protein)のような微小管修復を仲介する微小管関連タンパク質の局在は、生きた細胞における修復部位を同定するための代用品として役立つ。これらの部位は、高いコンフォメーションストレスを受けた微小管に沿って多数存在するが、軸索の直線微小管沿いにも存在し、そこではモーターを使った輸送が特に激しい。重要なことは、新しいGTP-チューブリン二量体が組み込まれると、微小管に沿ってレスキュー部位が生成され、その寿命が延びることである。このことは、ダイニンとキネシンのモーターが、それらが歩いている微小管を選択的に安定化させる引き金となり、微小管ネットワークにおける優先的なトラックの定義と細胞の極性の確立に役立っている可能性を提起する。